И. А. Мельников. Экосистемы морских льдов Антарктики: сравнительный анализ
УДК 551.46:54; 551.58
Введение
Зона морских льдов Антарктики представляет собой уникальное природное образование, подверженное значительным сезонным изменениям. В зимний период огромная площадь открытой воды вокруг континентальной Антарктики замерзает, образуя широкий пояс морских льдов площадью до 20 млн км2, а летом из-за таяния его площадь сокращается до 4 млн км2 (рис. 1). В результате таких ежегодных процессов образующийся морской ледовый покров оказывает колоссальное влияние на глобальную климатическую систему Земли, регулируя тепловые потоки между теплым Южным океаном и холодной атмосферой Антарктики. Зимой, когда температурный градиент между ними максимальный, потеря тепла в районах, занятых морскими льдами, может быть на два порядка меньше, чем в открытом от льда океане. Обладая высоким альбедо, морской лед со снежным покровом отражает большую часть падающей на его поверхность солнечной радиации, в то время как в открытом океане она интенсивно поглощается водной поверхностью.
![Рис. 1. Максимальная в сентябре (а) и минимальная в феврале (б) средняя месячная площадь морского ледникового покрова Антарктики по данным спутниковых наблюдений за период 1973-1976 гг. [Zwally et al., 1983]](pic/000066.jpg)
Рис. 1. Максимальная в сентябре (а) и минимальная в феврале (б) средняя месячная площадь морского ледникового покрова Антарктики по данным спутниковых наблюдений за период 1973-1976 гг. [Zwally et al., 1983]
Морской ледовый покров как физическая среда обладает более низкой соленостью по сравнению с морской водой. В период ледообразования накапливающийся во льду рассол выпадает в воду, увеличивая ее соленость и плотность, что в свою очередь стимулирует зимнюю конвекцию и способствует подъему с глубины к поверхности свежих вод, богатых питательными веществами. Весной, напротив, когда лед тает, накапливающаяся пресная вода способствует формированию на поверхности стабильного слоя пониженной солености. Влияние морского льда на взаимодействие между атмосферой и океаном зависит от распределения льда, его сплоченности и толщины, что в свою очередь влияет на такие гидрометеорологические параметры, как температура, скорость и направление ветра и течений, волнения и др. Таким образом, взаимодействия между океаном, льдом и атмосферой формируют сложную взаимосвязанную систему.
Морской ледовый покров Антарктики как биологическая среда представляет собой сложный природный комплекс, с которым биотопически связаны разнообразные биологические сообщества, начиная от одноклеточных ледовых водорослей и кончая тюленями и пингвинами, использующими морской лед как физическую платформу для размножения и защиты от хищников. Криль, трофически связанный с органической продукцией ледовых водорослей в зимний период и фитопланктоном в летний, является ключевым организмом в пищевой сети Антарктики.
В настоящее время изменения природной среды Антарктики достаточно хорошо изучены, особенно в районе Антарктического полуострова, где вследствие потепления за последние 40 лет наблюдаются заметное увеличение площади открытой от льда воды (в форме разводий и полыней) и соответственно сокращение площади морского ледового покрова и продолжительности его существования, а также разрушение покровных ледников [Doake, Vaughan, 1991; Zwally, 1991; Gammie, 1995; Vaughan, Doake, 1996; Rott et al., 1996; Hewitt, 1997]. Вследствие потепления в данном районе зарегистрировано увеличение численности сальпы Salpa thomsoni на фоне заметного уменьшения численности и биомассы эуфаузииды Euphausia superba [Loeb et al., 1997]. Эти два вида, трофически конкурирующие в морской экосистеме Антарктики, являются надежным индикатором изменений, наблюдаемых в настоящее время в этом районе. Вместе с тем, несмотря на явную очевидность глобальных изменений морской среды в Арктике [Johannessen et al., 1995; Cavaliere et al., 1997; Carmarck et al., 1997; Serreze, Maslanik, 1997; McPhee et al., 1998; Morrison et al., 1998; Vinnikov et al., 1999] и их влияние на состав, структуру и динамику экосистем морского льда и верхнего океана [Мельников и др., 2000; Melnikov et al., 1998; Melnikov, 2000; Melnikov et al., 2001], в Антарктике нет очевидных свидетельств крупномасштабных изменений состава биологических сообществ морской среды под влиянием глобального потепления, кроме района Антарктического полуострова и районов субантарктических островов [Smith, Steenkamp, 1990; Smith, 1991]. Однако при устойчивом сокращения площади морских льдов можно предположить существенную перестройку в составе и структуре экосистемы морской Антарктики, что приведет к драматическим изменениям в ее пищевой сети, а следовательно, к глобальному изменению потоков углерода в этом важном районе Земли.
В статье не ставится целью рассмотрение сценария функционирования современной экосистемы зоны морских антарктических льдов под действием климатического фактора. Напротив, автор полагает, что для рассмотрения такого (или таких) сценария необходимы фундаментальные знания о составе, структуре и функционировании морских экосистем вообще и ледовой экосистемы в частности. Понимание природы наблюдаемых изменений в Антарктике позволит в дальнейшем прогнозировать поведение ее биологических систем под воздействием природных или антропогенных факторов среды. Цель нашей работы - рассмотрение биологических характеристик основных элементов ледового покрова морской Антарктики - сезонных льдов, формирующихся ежегодно над глубоким океаном вокруг континента, и припайных льдов, образующихся в мелководной прибрежной зоне в зимний период, т. е. здесь будут рассматриваться раздельно экосистемы пелагических и прибрежных морских льдов. Нам представляется важным и своевременным дать обзор основных экосистем зоны морских льдов Антарктики в период, когда она начинает испытывать первые признаки глобального потепления. В основу обзора положены материалы, полученные главным образом в период наблюдений на континентальной польской станции им. Г. Арцтовского в 1987-1988 гг. [Мельников, 1993] и в период дрейфа российско-американской станции в западной части моря Уэдделла в феврале-июне 1992 г. [Мельников, 1995], а также литературные данные.
В статье все термины и понятия используются в соответствии с общепринятой номенклатурой, принятой для ледового покрова [Зубов, 1945; Волков, Трешников, 1968] и ледовых организмов [Усачев, 1949; Мельников, 1989; Horner et al., 1988, 1992]. Прежде всего необходимо отметить, что в понятие "ледовая флора и фауна" входят все водоросли и животные, связанные со льдом независимо от их биотопической принадлежности, т. е. это понятие объединяет весь флоро-фаунистический комплекс морского льда. В зависимости от биотопической принадлежности организмов в данном исследовании всю ледовую флору и фауну мы разделяем на три экологические группы: 1) население нижней (морской) поверхности; 2) население толщи льда и 3) население верхней (воздушной) поверхности льда.
Морские льды пелагиали
Основная масса льда, формирующегося зимой на открытой воде после летнего таяния, распространяется широким поясом вокруг всего континента. Это дрейфующий паковый лед, занимающий район с наибольшими изменениями ледовых условий. Он содержит различные типы льдов, различающиеся по возрасту, толщине и сплоченности, его дрейф определяется ветром и течениями. Вследствие существующих напряжений между льдинами часто образуются пространства открытой воды, которая замерзает, поэтому в одном и том же месте всегда можно встретить лед на различных стадиях развития.
Площадь ледового покрова в период максимального развития превосходит площадь континентальной Антарктики. Положение северной границы кромки ледового покрова изменяется год от года. Циклонические течения (например, в морях Росса и Уэдделла) существенно влияют на распределение и дрейф льдов в Южном океане. Так, паковый лед, связанный с круговоротом моря Уэдделла, распространяется далеко на север, дальше, чем в любом другом районе Антарктики (до 2200 км от континента). В восточной Антарктике (например, в секторе между 120 и 135° з. д.) паковый лед напротив, может существовать лишь на расстоянии нескольких сотен километров от берега. Это следствие того, что береговая линия в этом районе расположена севернее, чем в любом другом месте Антарктики (за исключением района вблизи Антарктического п-ова), поэтому здесь теплее, что способствует более интенсивному таянию. За короткое лето положение кромки может существенно меняться со скоростью несколько десятков километров в день из-за характерных локальных синоптических условий, связанных с изменчивостью направления ветра.
В отличие от Арктического бассейна, где морские льды как бы зажаты между континентами, морские льды Антарктики, напротив, находятся за пределами континета, поэтому ее ледовый покров очень динамичен и испытывает циклические периоды конвергенции и дивергенции в результате действия ветра и течений. К северу от антарктической дивергенции пак движется в основном с запада на восток, с преобладающим направлением дрейфа льда на север. Вследствие такой циркуляции и сезонной изменчивости лед активно выносится за пределы антарктического циркумполярного течения, где разрушается под действием волн и активного таяния в более теплых водах, и лишь небольшая по площади часть ледового покрова сохраняется более чем один сезон, как правило, в небольших заливах и бухтах или в западной части моря Уэдделла и в море Росса, где встречаются остаточные (двухлетние) льды.
Дрейф льда - важная характеристика морского ледового покрова пелагиали Южного океана. Генеральный дрейф - дивергентный, но часто случается и конвергентный, что становится причиной разлома ледовых полей. Дивергентный дрейф поддерживается южным ветром, что заставляет лед двигаться в северном направлении, увеличивая сплоченность и толщину льда за счет сжатия и деформации ледовых полей. Характерное направление дрейфа льда в Восточной Антарктике - западное (вблизи континента) и восточное у северной границы Антарктической дивергенции. Данные спутниковых наблюдений за дрейфом 45 буев за период с 1985 по 1997 г. в Восточной Антарктике показали, что средняя скорость дрейфа льда в западном направлении составляет 0,22 м/с (19,4 км/день), а в восточном - 0,17 м/с (14,9 км/день) [Worby, 1999]. Скорость дрейфа меняется в пространстве и времени и иногда достигает 0,9 м/с (78 км/день). В Западной Антарктике, например, в период дрейфа станции Уэдделл-1 максимальная скорость дрейфа составила в северном направлении 25 км/день (24 мая), а минимальная 0 5 км/день (27 февраля) [Gordon, Lukin, 1992].
Первые кристаллы льда образуются на открытой поверхности воды, когда ее температура снижается до -1,8 °С. Образующиеся кристаллы придают воде "маслянистые" свойства, из-за чего такой лед называют сало (рис 2 а) Дальнейший рост кристаллов зависит от синоптических условии: при неспокойной погоде (ветре, волнении) на поверхности воды образуется так называемый блинчатый лед (рис. 2, б), пластины которого могут достигать размеров от нескольких сантиметров до десятков метров в диаметре. При спокойных условиях и кристаллы, и сало могут смерзаться, образуя вязкую массу, называемую шугой (рис. 2, в), или формировать тонкую вязкую пленку - нилас - толщиной до 10 см. Чаще всего этот лед ломается, пластины надвигаются одна на другую и смерзаются, механически увеличивая толщину льда (рис. 2, г). Скорость роста льда на открытой воде за первые 24 часа составляет 0,38 см/ч (9 см/сут) и затем снижается до 0,03 см/сут, достигая толщины 28 см на 8-й день [Melnikov, 1995]. Хотя новообразовавшийся лед растет быстро, однако на начальной стадии формирования его биогеохимические ингредиенты меняются незначительно. При исследовании солености, концентрации минерального кремния и хлорофилла а в новообразующемся льду на станции Уэдделл-1 в мае 1992 г. [Melnikov, 1995] мы наблюдали активную динамику солености и кремния в первые двое суток (рис. 3, а, б), когда лед еще тонок (около 12-15 см) и поэтому сильно охлажден, т. е. это время максимального выхода солей из льда в воду. В первые сутки концентрация хлорофилла а составляла всего 0,01 мкг/л (рис. 4), и в пробах были определены лишь отдельные клетки диатомовой водоросли Fragilariopsis cylindrus. Начало заметного роста концентрации хлорофилла а было отмечено на третьи сутки, когда в нижних слоях, где концентрируются одноклеточные водоросли, условия среды стали благоприятными для развития клеток: их биомасса (по содержанию хлорофилла а) стала плавно расти достигнув значения 0,23 мкг/л на восьмой день (см. рис. 4).
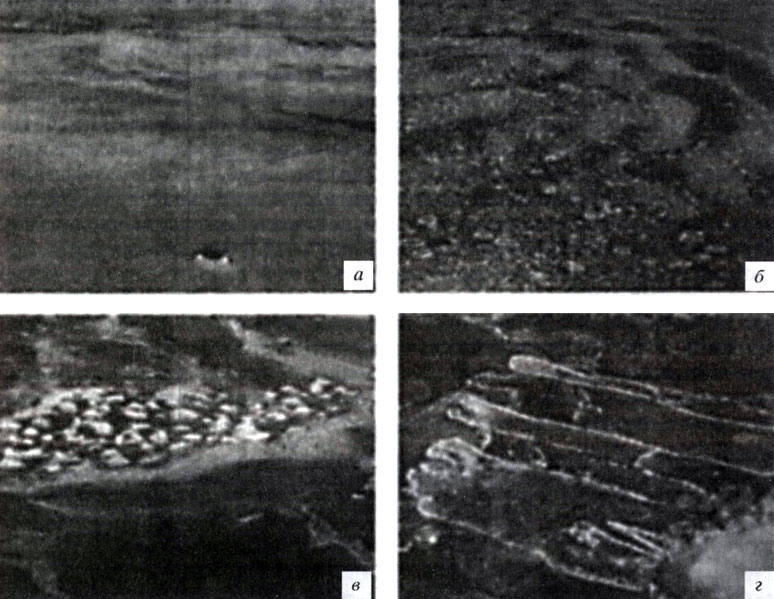
Рис. 2. Начальные формы молодого льда, формирующегося на открытой воде в пелагиали Антарктики: а - ледяное сало; б - блинчатый лед; в - шуга; г - нилас. Фотографии публикуются с разрешения Ian Allison (Australian Antarctic Division). Объяснения см. в тексте
![Рис. 3. Динамика солености (5) и минерального кремния (Si) на начальной стадии формирования льда на открытой воде в разводье в 24-часовом эксперименте 20-21 мая (а) и 8-дневном эксперименте 21-28 мая (б); ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]](pic/000068.jpg)
Рис. 3. Динамика солености (5) и минерального кремния (Si) на начальной стадии формирования льда на открытой воде в разводье в 24-часовом эксперименте 20-21 мая (а) и 8-дневном эксперименте 21-28 мая (б); ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]
![Рис. 4. Хлорофилл а в 24-часовом (20-21 мая) и 8-дневном (20-28 мая) эксперименте по формированию льда на открытой воде в разводье; ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]](pic/000069.jpg)
Рис. 4. Хлорофилл а в 24-часовом (20-21 мая) и 8-дневном (20-28 мая) эксперименте по формированию льда на открытой воде в разводье; ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]
При дальнейшем росте лед постепенно заселяется микрофлорой, которая проникает из воды в нижние слои по капиллярной системе. Поскольку температурные условия в слоях, прилегающих к границе раздела вода-лед, близки к таковым в воде, то клетки начинают активно развиваться в этих слоях, а при дальнейшем росте льда и увеличении его толщины они развиваются и в его толще (рис. 5). На рис. 6 и 7 представлены данные по динамике солености и концентрации минерального кремния и хлорофилла а в однолетнем и двухлетнем льду. Максимальное развитие водорослей в однолетнем льду наблюдается в нижнем контактирующем с водой слое, где их биомасса (в величинах концентрации хлорофилла а) достигает 30-35 мкг/л, что на два порядка выше, чем в воде подо льдом (таблица). Напротив, в старом (двухлетнем) льду максимальное развитие водорослей наблюдается в верхнем слое - на границе раздела лед-снег, где величина биомассы водорослей (по хлорофиллу а) составляет 45-40 мкг/л. В обоих случаях было выявлено 88 и 89 видов диатомовых водорослей соответственно. Два вида - Fragilariopsis cylindus и Archaemonas areolata - (более 80% численности от общего списка водорослей) доминировали в зимний период в двухлетнем льду, a f. cylindus (около 90% численности) в однолетнем льду. Диатомовые Chaetoceros deflandrei, Ch. debilis, Ch. socialis доминировали в обоих типах льда в конце зимы, a Corethron criophilum и Eucampia antarctica - в начале весны.
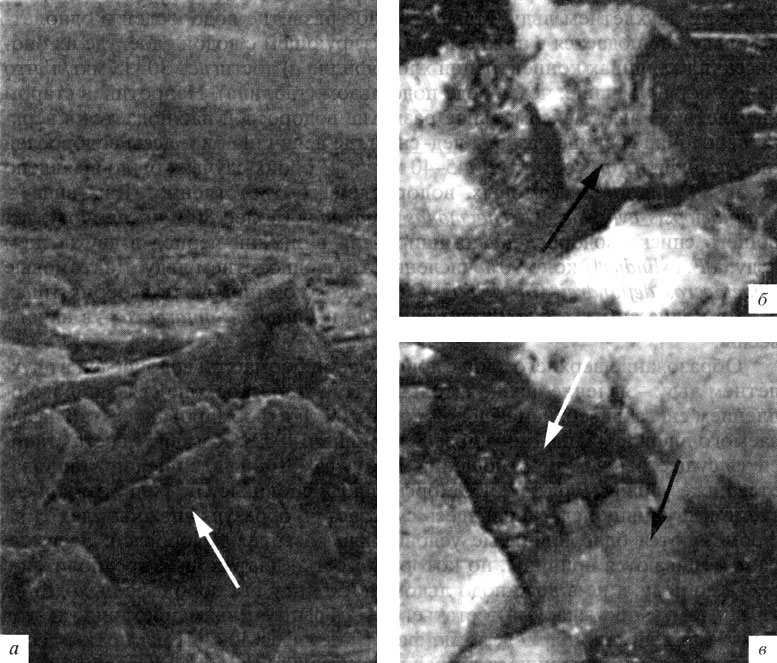
Рис 5 Положение слоев диатомовых водорослей в морском льду в пелагиали Антарктики: а - в толще льда; б - на границе раздела лед-снег; в - на границе раздела лед-вода. Фотографии предоставлены S. Nicol (Australian Antarctic Division)
![Рис. 6. Динамика солености (5), минерального кремния (Si) и хлорофилла а (СЫ а) в верхнем (1), среднем (2) и нижнем (3) слоях однолетнего льда в марте-июне 1992 г.; ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]](pic/000071.jpg)
Рис. 6. Динамика солености (5), минерального кремния (Si) и хлорофилла а (СЫ а) в верхнем (1), среднем (2) и нижнем (3) слоях однолетнего льда в марте-июне 1992 г.; ледовая станция Уэдделл-1, 1992 [Melnikov, 1995]
![Рис. 7. Динамика солености (S), минерального кремния (Si) и хлорофилла а (СЫ а) в верхнем (7), среднем (2) и нижним (3) слоях двухлетнего льда, март-июнь 1992 г.; ледовая станция Уэд-делл-1 [Melnikov, 1995]](pic/000072.jpg)
Рис. 7. Динамика солености (S), минерального кремния (Si) и хлорофилла а (СЫ а) в верхнем (7), среднем (2) и нижним (3) слоях двухлетнего льда, март-июнь 1992 г.; ледовая станция Уэд-делл-1 [Melnikov, 1995]
Образование верхнего слоя с повышенной биомассой водорослей в двухлетнем льду - явление исключительно антарктическое, оно связано с накоплением снега на верхней ледовой поверхности и формированием так называемого инфильтрационного льда [Буйницкий, 1973]. Когда вес снега становится существенным по отношению к весу самого льда, то лед притаплива-ется ниже уровня моря так, что морская вода проникает по капиллярной системе на границу лед-снег и здесь смерзается, образуя "снежный лед", где формируются благоприятные условия для развития водорослей. Здесь не рассматриваются причины, по которым клетки диатомовых предпочитают расти во льду, а не в воде подо льдом, поскольку эта проблема выходит за рамки темы, обсуждаемой в статье. Мы обращаем внимание лишь на тот факт, что в зимний период, когда фотосинтез фитопланктона практически отсутствует, развивающиеся во льду диатомовые водоросли создают органическое вещество, количество которого многократно превышает его концентрацию в воде подо льдом [Melnikov, 1998]. Это органическое вещество является пищевым депо для криопелагической фауны [Menshenina, Melnikov, 1995], криля [Melnikov, Spiridonov, 1996], зоопланктона [Pasternak, 1995] и рыб [Evseenko, 1994], биотопически связанных с морским льдом пелагиали Антарктики.
| МОРСКОЙ ЛЕД ПЕЛАГИАЛИ | ||||||
|---|---|---|---|---|---|---|
| Тип льда | Chl a, мкг/л | РОС, мкг/л | Si, мкг/л | Соотношение лед/вода | ||
| Chl a | РОС | Si | ||||
| Молодой | 14,7 (n=40) | 1602 (n=40) | 335 (n=40) | 245 | 21 | 0,17 |
| Однолетний | 7,6 (n=51) | 889 (n=51) | 207 (n=51) | 122 | 12 | 0,11 |
| Двухлетний | 6,1 (n=78) | 714 (n=78) | 99 (n=78) | 98 | 9 | 0,05 |
| Морская вода | 0,06 (n=9) | 76 (n=9) | 1928 (n=9) | - | - | - |
| ПРИБРЕЖНЫЙ МОРСКОЙ ЛЕД | ||||||
| Молодой | 1,1 (n=9) | 243 (n=28) | 36 (n=27) | 1,69 | 2,67 | 0,44 |
| Припайный | 69 (n=17) | 1565 (n=7) | 16 (n=7) | 106 | 17,2 | 0,19 |
| Якорный | 17 (n=44) | 1850 (n=44) | 17 (n=43) | 26 | 20,3 | 0,21 |
| Морская вода | 0,65 (n=14) | 97 (n=16) | 81 (n=18) | - | - | - |
Льды прибрежной зоны
В прибрежной зоне доминируют два типа льда: аллохтонный, т. е. местный, образующийся на открытой воде залива, и автохтонный, т. е. образующийся на других акваториях и вносимый извне вследствие взаимодействия течений и ветра. К первому типу относятся припайный и собственно береговой льды, которые, как правило, неподвижны (по крайней мере, до момента взлома припая). Ко второму типу относятся дрейфующие льды, формирующиеся на открытой воде. И те, и другие включают как глетчерные (айсберги, обломки покровных ледников), так и морские льды, различающиеся по происхождению, возрасту и подвижности (рис. 8). Так, ледовые поля, образующиеся на открытой воде, взламываются на мелкие льдины, которые могут далее переноситься течениями в прибрежную зону, где в отлив остаются на дне, образуя так называемый якорный лед (рис. 9). Хотя количество такого льда не превышало 10% от общей площади (Адмиральский залив, о-в Кинг-Джордж, зима 1987 г.), воздействие такого заякоренного льда на экосистему литорали чрезвычайно велико. У нас нет прямых наблюдений за последствием таких воздействий в Антарктике, поэтому мы судим о них по нашим наблюдениям в Белом море (неопубликованные собственные данные), где климатические условия схожи с таковыми у Антарктического п-ова. Так, в прибрежной зоне Кандалакшского залива, где приливо-отливные амплитуды схожи с амплитудами Адмиральского залива и достигают 2 м, бентос каменистой литорали теряет за зиму до 90% фауны литорин и баля-нусов (мелкие моллюски и усоногие раки) и флоры фукоидов (бурые водоросли) вследствие механического воздействия льда на дно литорали. В Антарктике береговая линия (вплоть до глубины 15 м) подвергается воздействию льда (припайного, пакового, якорного). Айсберги всех размеров могут разрушать бентос. Когда они вносятся течением, то их подводные (донные) части вспахивают мягкие грунты, что приводит к разрушению бентоса. На восстановление нарушеных сообществ требуются годы. Кроме того, на дне образуется внутриводный лед толщиной от 2-5 мм до 15 см (см. рис. 9, д). Такой лед также может нарушать бентос. Известен случай, когда внутри-водный лед поднял к поверхности 25 кг бентосных животных, вмерзших в лед при его образовании [Dayton et al., 1969].
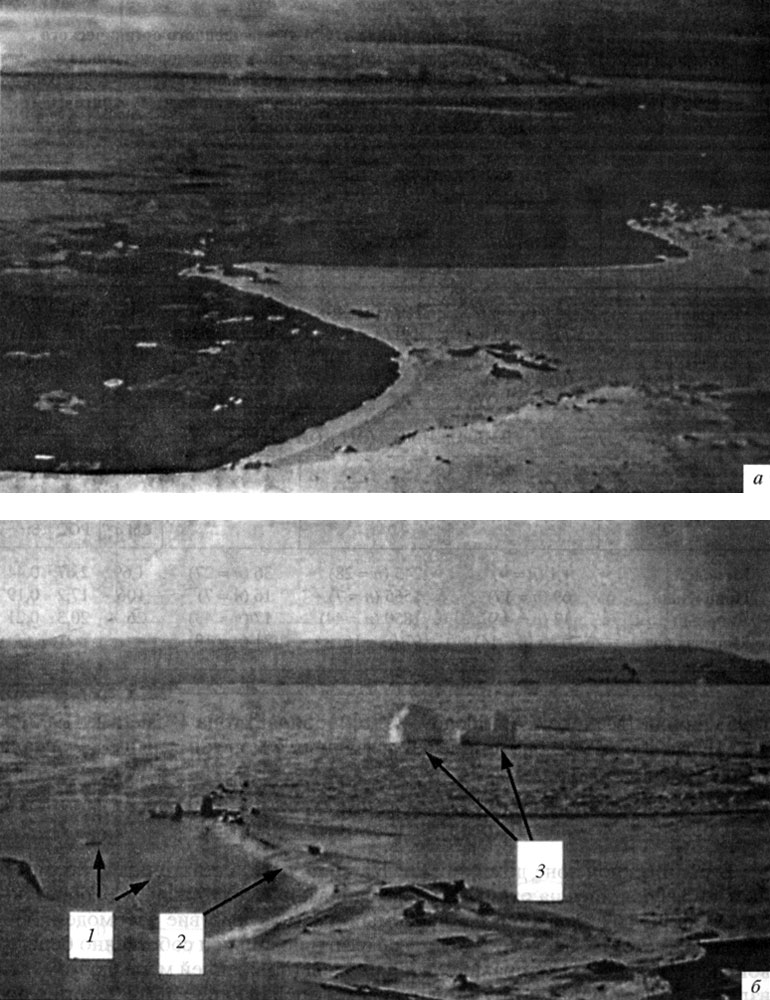
Рис. 8. Адмиральский залив у польской антарктической станции им. Г. Арцтовского (о-в Кинг-Джордж): а - свободный от льда в июне; б-с припайным льдом, образовавшимся в августе 1988 г. Стрелками показано положение заякоренных льдин в приливо-отливной части залива (7), берегового льда (2) и двух заякоренных айсбергов в глубоководной части залива (3)
Припайный лед образуется в конце мая, ранее, чем замерзает сам залив, и сохраняется до конца сентября. Физические факторы, такие как орография берега, приливо-отливные течения и ветер, определяют начало формирования припая и время его существования. В районе о-ва Анверс (Антарктический п-ов, 64° ю. ш.) лед существует более шести месяцев в году; в районе залива Мак-Мердо (море Росса, 77° ю. ш.) - более одиннадцати месяцев, а у о-ва Сигни (Южные Оркнейские о-ва, 60° ю. ш.) - не более одного-двух месяцев [Worby, 1999]. За 15 лет наблюдений (1957-1973 гг.) продолжительность существования льда у о-ва Сигни изменялась от 70 до 241 дней (в среднем 144 дня). Зимой 1987 г. Адмиральский залив (о-в Кинг-Джордж) замерзал в июне несколько раз, но всякий раз сильный ветер и волнение взламывали лед и выносили его из залива. В начале июля залив замерз, и лед продержался в течение 46 дней, достигнув толщины 46 см в центре залива и 73 см на его периферии. С 18 на 19 августа, после сильного ветра, лед взломало и вынесло в пролив Брансфильд, и только в небольших заливчиках лед сохранился до конца сентября. В летнее время в заливе отмечены обломки морского льда, вносимого в залив из пролива Брансфильд, или отколовшиеся от ледников, окаймляющих залив, а также от вносимых в залив извне айсбергов (см. рис. 8). Появление якорного льда определяется взаимодействием приливо-отливных течений и ветра: в высокую воду ветер пригоняет льдины к берегу, а когда начинается отлив и вода уходит, то лед, как бы обсыхает и остается на мели, т. е. заякоривается (см. рис. 9, а, б, в, г).
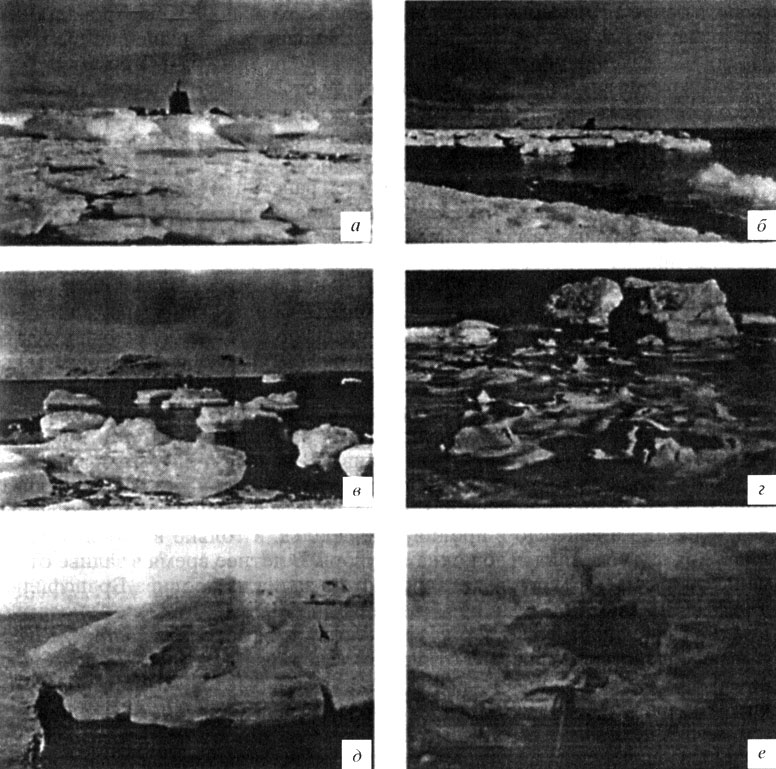
Рис. 9. Типы морских льдов в прибрежной зоне Адмиральского залива у континентальной польской антарктической станции им. Г. Арцтовского на о-ве Кинг-Джордж зимой 1988 г.: а, б, в - заякоренные льды в приливо-отливной зоне; г - донный (обсохший) лед в отливе; д - льдины однолетнего льда, внесенного в залив из пролива Брансфильда; е - береговой лед. На последних двух снимках четко видны слои с вкраплениями ледовых водорослей
Образование льда на открытой воде в припайной зоне идет аналогично процессам, происходящим в пелагиали, однако на мелководье, в приливо-отливной зоне, имеются свои характерные черты ледообразования. Механизм нарастания сверху типичен для берегового льда. В приливо-отливной зоне лед "наплывает" на берег, образуя хаотичные торосистые образования, которые в результате многократного воздействия приливов и отливов "заяко-риваются", создавая своеобразные плотины (см. рис. 8, б). Когда вода достигает максимальной береговой отметки во время прилива, наступает момент, когда она растекается (наплывает) сверху по льду. При сильных морозах вода замерзает на верхней ледовой поверхности, образуя своеобразные наледи. Их мощность зависит от морфологии берега, характера дна, интенсивности приливов-отливов и т. д. Именно в момент прилива, когда вода заливает верхнюю (снежную) поверхность, образуя снежницы, здесь развивается фитопланктон, "цветение" которого на снежно-ледяной поверхности наблюдается в течение всей зимы практически постоянно.
На основании круглогодичных наблюдений в Адмиральском заливе на о-ве Кинг-Джордж в 1987-1988 гг. было показано, что биомасса водорослей (в величинах концентрации хлорофилла а) на 2-3 порядка превосходит биомассу водорослей в воде под льдом (см. таблицу). Соотношения между величинами взвешенного органического углерода и хлорофилла а составили 220 в молодых формах льда, 109 - у якорного льда и 23 - в береговом льду. Максимальные концентрации хлорофилла а (210 мкг/л) и взвешенного органического углерода (около 6000 мкг/л) были измерены в береговом льду. Здесь было зарегистрировано 119 видов водорослей, среди которых Navicula gelida var. Parvula, Nitzschia lecontei, N. prologatoides, N. barbieri, Fragilariopsis cylindrus и F. curta (95% от общей численности водорослей) доминировали в заякоренных льдах, a Navicula gelida, Nitzschia lecontei, N. barbieri, Corethron criophilum, Fragilariopsis cylindrus и F. curta (90%) - в припайном льду.
Важно отметить, что такое высокое содержание органического вещества во льдах прибрежной зоны поддерживается за счет высокой биологической активности ледовых водорослей, и, несомненно, их продукция является дополнительным энергетическим депо для биологических сообществ литорали и сублиторали, биотопически связанных с зоной припайных льдов в зимний период.
Благодарность. Работа выполнена в рамках Госконтракта РФ по реализации подпрограммы "Изучение и исследование Антарктики" ФЦП "Мировой океан" проекта № 17 "Оценка экологии зоны морских льдов Антарктики" ("Криаль").
Литература
Буйницкий В. Х. Морские льды и айсберги Антарктики. Л.: Изд-во ЛГУ, 1973. 255 с. Волков Н.А., Трешников А.Ф. О новой международной номенклатуре морских льдов//Пробл. Арктики и Антарктики. 1968. № 32. С. 136-143.
Зубов Н. Н. Льды Арктики. М.; Л.: Изд. Главсевморпути, 1945. 360 с.
Мельников И. А. Экосистема арктического морского льда. М.: ИОАН, 1989. 192 с.
Мельников И. А. Хлорофилл "а", взвешенный органический углерод и минеральный кремний в морском льду залива Адмиралти (о. Кинг-Джордж, Зап.Антарктика)//Пелагические экосистемы Южного океана. М: Наука, 1993. С. 83-91.
Мельников И. А. Ледовая дрейфующая станция "Уэдделл-1" (Антарктика)//Океанология. 1995. Т. 35, №2. С. 310-313.
Мельников И. А., Колосова Е. Г., Житина Л. С. Реакция морских экосистем на глобальные изменения в Арктике: верхний океан//Труды Арктического регионального центра. Владивосток: Дальнаука, 2000. С. 69-80.
Усачев П. И. Микрофлора полярных льдов//Тр. ИОАН. 1949. Т. 3. С. 216-259.
Carmack E.C., Aagaard К., Swift J.H. et al. Changes in temperature and tracer distributions within the Arctic Ocean: Results from the 1994 Arctic ocean section//Deep-Sea Res. 1997. Vol. 44, N 8. P. 1487-1502.
Cavaliere D. J., Gloersen P., Parkinson C. L. et al. Observed hemispheric asymmetry in global sea ice changes//Science. 1997. Vol. 278. P. 1104-1106.
Dayton P. K., Robilliard G. A., De Vries AL. Anchor ice formation in McMurdo Sound, Antarctica, and its biological effects//Science. 1969. Vol. 163. P. 273-274.
Doake C. S. M., Vaughan D. G. Rapid disintegration of the World Ice Shelf in response to atmospheric warming//Nature. 1991. Vol. 350. P. 328-330.
Evseenko S. A. Fish larvae and juveniles from the western Weddell Sea//Proc. SCAR VI Biol. Symp., Venice, Italy, 30 May-3 June, 1994. Venice, 1994. P. 87.
Gammie F. Breakway iceberg due to warming//Nature. 1995. Vol. 274. P. 108.
Gordon, A. L., Lukin V. V. Ice Station Weddell N 1//Antarct. J.US. 1992. Vol. 27. P. 97-99.
Hewitt R. P. Areal and seasonal xtent of sea-ice cover off the northwestern side of the Antarctic Peninsula: 1979-1996//CCAMLR Science. 1997. Vol. 4. P. 65-73.
Horner R. A., Ackley S. F., Dieckmann G. S. et al. Ecology of sea ice biota. 1. Habitat, terminology, and methodology//Polar Biol. 1992. Vol. 12. P. 417-427.
Horner R. A., Syvertsen E. E., Thomas D.P., Lange C. Proposed terminology and reporting units for sea ice algal assemblages//Ibid. 1988. Vol. 8. P. 249-253.
Johannessen O. M., Miles M., Bjorgo E. The Arctic's shrinking sea ice//Nature. 1995. Vol. 376. P. 126-127.
McPhee M., Stanton T. P., Morison J. H., Martinson D.G. Freshening of the upper ocean in the Arctic: Is perennial sea ice disappearing?//Geophys. Res. Lett. 1998. Vol. 25. P. 1729-1732.
Melnikov L. A. An in situ experimental study of young sea ice formation on Antarctic lead//J. Geophys. Res. 1995. Vol. 100, N C3. P. 4673-4680.
Melnikov I. A. Winter production of sea ice algae in the western Weddell Sea//J. Mar. Systems. 1998. Vol. 17, N 1/4. P. 195-206.
Melnikov L. A. The Arctic sea ice ecosystem and global warming//Impacts of changes in sea ice and other environmental parameters in the Arctic: Final report of the Marine Mammal Commission Workshop, Girdwood, Alaska, 15-17 Febr., 2000 / Ed. H.P. Huntington. Bethesda (Md.): Mar. Mammal Commiss., 2000. P. 94-110.
Melnikov I A., Sherr В., Wheeler P., Welch H. Preliminary biological and chemical oceanographic evidence for a long-term warming trend in the Arctic Ocean (current materials of the SHEB A Ice Camp, Beaufort Sea)//Proc. Arctic Change Workshop, Seattle, Univ. of Wash., June, 1997. Seattle (Wash.), 1998. Rep. 8. P. 60.
Melnikov I. A., Spiridonov V. V. Antarctic krill, Euphausia superba Dana (Crustacea Euphausiacea) under perennial sea ice in the western Weddell Sea//Antarct. Sci. 1996. Vol. 8, N 4. P. 323-329.
Melnikov I. A., Zhitina L. S., Kolosova E.G. Arctic sea ice biological signal of global warming//Proc. NIPR Symp. Polar Biol. 2001. N 13.
Menshenina L. L., Melnikov I. A. Under-ice zooplankton of the western Weddell sea//Ibid. 1995. N 8. P. 126-138.
Morison J., Steele M., Anderson R. Hydrography of the upper Arctic Ocean measured from the nuclear submarine USS Pargo//Deep-Sea Res. 1998. Vol. 45, pt 1. P. 15-38.
Pasternak A. F. Cut content and feeding rhythm in dominant copepods in the ice-covered Weddell Sea, March 1992//Polar Biol. 1995. Vol. 15. P. 583-586.
RottH., Skvarca P., Nagler T. Rapid collapse of northern Larsen ice shelf, Antarctica // Science. 1996. Vol. 271. P. 788-792.
Serreze M. C., Maslanik J. A. Polar processes in global climate, 13-15 Nov., 1996, Cancum, Mexico. Boston: Amer. Meteorol. Soc, 1997. P. 110-112.
Smith V. R. Climate change and its ecological consequences at Mariot and Prince Edward Islands//S. Afr. Antarct. Res. 1991. Vol. 21. P. 223-224.
Smith V. R., Steenkamp M. Climate change and its ecological implications at a subantarctic island//Oecologia. 1990. Vol. 85. P. 14-24.
Vaughan D. G., Doake C. S. M. Recent atmospheric warming and retreat of ice shelves on the Antarctic Peninsula//Nature. 1996. Vol. 379. P. 328-331.
Vinnikov K. Y., Robok A., Stouffer R. et al. Global warming and northern hemisphere sea ice extent//Science. 1999. Vol. 286, N 5446. P. 1934-1937.
Worby A. P. Observing Antarctic sea ice: A practical guide for conducting sea ice observations from vessels operating in the Antarctic pack ice. A CD-ROM produced from the Antarctic Sea Ice Processes and Climate (ASPeCT) program of the SCAR Global Change and the Antarctic (GLOCHANT) Program. Hobart (Australia), 1999.
Zwally H. J. Break-up of Antarctic ice//Nature. 1991. Vol. 350. P. 274.
Zwally H. J., Comiso J. C., Parkinson C. L. et al. Antarctic sea ice, 1973-1976: Satellite passive-microwave observations. Wash. (D.C.), 1983. P. 206 (NASA SP-459).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://antarctic.su/ 'Арктика и Антарктика'